

推荐产品





公司新闻/正文
安捷伦 Seahorse XF 分析仪和 XF T 细胞代谢分析试剂盒评估 NK 细胞的生物能量代谢
295 人阅读发布时间:2025-08-20 11:02
摘要
代谢已经逐渐被认为是免疫细胞命运和功能的关键驱动因素。NK 细胞需要通过糖酵解和 OXPHOS(氧化磷酸化)来实现 NK 细胞活化、增殖和效应功能。此外,最近的研究报道了使用基因工程 NK 细胞 (CAR-NK) 进行癌症治疗的潜在益处,并且早期临床研究也获得了令人鼓舞的初步数据。
调节 NK 细胞代谢是改善基于 NK 细胞的癌症免疫疗法疗效的潜在策略。安捷伦 Seahorse XF NK 细胞代谢分析检测使用安捷伦 Seahorse XF 分析仪和安捷伦 Seahorse XF T 细胞代谢分析试剂盒实时、全面评估 NK 细胞代谢。该方法可以同时测量 NK 细胞群中的糖酵解和线粒体活性以及生物能量代谢能力。XF T 细胞代谢分析试剂盒使用了一种改进的解偶联剂 (BAM15),能够更可靠、准确地测量 NK 细胞线粒体生物能量代谢能力,大大降低了试剂优化需求。本应用简报概述了使用该检测方法在细胞扩增过程中以及细胞因子刺激后监测和分析 NK 细胞代谢。此外,还展示了在设计和开发 NK 细胞治疗产品的最佳生产工艺时的潜在用途。
前言
自然杀伤 (NK) 细胞是先天免疫反应的关键参与细胞。它们可以识别细胞表面的特异性抗原肽,并与感染了病毒、细菌或由致癌基因转化的细胞上表达的配体结合。它们在抗病毒反应和支持抗肿瘤活性方面也发挥着关键作用。最近,研究人员认识到,NK 细胞可以促进 T 细胞反应的形成,在适应性反应中发挥作用。此外,最近的研究报道了使用基因工程 NK 细胞 (CAR-NK) 进行癌症治疗的潜在益处,并且早期临床研究也获得了令人鼓舞的初步数据。
不论 NK 细胞来源(脐带血、iPSC、外周血)如何,制备基因工程 NK 细胞群都需要在将其输注回患者体内之前进行大量体外扩增2。研究人员普遍认为,新鲜分离的 NK 细胞的细胞杀伤能力较低。NK 细胞扩增可采用不同的扩增系统(包括培养设备、培养基和细胞因子组成以及与饲养细胞系共培养),从而可以得到具有不同属性和体内疗效的细胞产品。此外,通过与不同的细胞因子组合(如 IL-12、IL-15 和 IL-18)孵育在体外“启动” NK 细胞,可诱导表型变化,被称为细胞因子诱导的记忆样 NK 细胞。这些“记忆样” NK 细胞在体内对肿瘤产生的反应更强烈3。
正如其他免疫细胞(如 T 细胞)所广泛证明的那样,细胞代谢在支持和调节 NK 细胞功能方面起着重要作用。代谢缺陷会导致产生功能失调的 NK 细胞1。通过优化扩增条件,可提高 NK 细胞的代谢适应性,从而增强体内抗肿瘤功能。
本应用简报介绍了使用安捷伦 XF NK 细胞代谢分析检测,同时可靠地测量 NK 细胞群的糖酵解和线粒体活性以及生物能量代谢能力。这些测量结果提供了 NK 细胞代谢谱的全面表征。这项研究采用一种检测方法即可监测和优化扩增的 NK 细胞群的代谢适应性,并筛选能够增强 NK 细胞治疗产品抗肿瘤功能的条件。
实验部分
小鼠 NK 细胞分离
使用“EasySep Mouse NK Cell Isolation Kit”,根据制造商的方案(货号 19855;STEMCELL Technologies,Vancouver,Canada),从来自 9 周龄未免疫 C57BL/6 小鼠(定制货号 CF-1201;Hooke Laboratories, LLC;Lawrence,MA)的 5 个新鲜脾脏中分离小鼠 NK 细胞。通过对小鼠 NK 细胞进行细胞表面标志物染色,确定 CD3-CD49+ 和 CD45+ 细胞群的百分比,从而确认分离出的小鼠 NK 细胞的质量。使用供应商推荐的抗体浓度对细胞进行染色。使用 FACS 缓冲液(DPBS,5% FBS,0.1% NaN3)制备 TruStain FcX PLUS(抗小鼠 CD16/32)抗体(货号 156603;BioLegend,San Diego,CA),并在冰上孵育 10 分钟。加入表面标志物抗体混合液,在冰上孵育 20 分钟,并在 FACS 缓冲液中清洗 3 次。3 次清洗完成后,将细胞重悬于 100 ng/mL DAPI 中,并使用 Agilent NovoCyte Advanteon 流式细胞仪搭配 Agilent NovoExpress 软件进行分析。所用的 BioLegend 抗体如下:抗小鼠 CD3- FITC(克隆号 17A2)(货号 100204)、抗小鼠CD45- PE 抗体(货号 103105)和抗小鼠 CD49b- APC(克隆号 DX5)(货号 108909)。
人 NK 细胞培养和扩增
根据制造商的说明解冻人外周血 NK 细胞(货号 0036;STEMCELL Technologies,Vancouver,Canada),并以300 × g 离心 10 分钟。然后以 1 × 106 个细胞/mL 将其重悬于 ImmunoCult NK 细胞基础培养基(货号 100-0712;STEMCELL Technologies)中,并置于 25 mL 培养瓶中在 37 °C、5% CO2 条件下过夜培养。在第 0 天使用细胞进行检测,或按照 ImmunoCult NK 扩增试剂盒(货号 100-0711;STEMCELL Technologies)的制造商方案进行扩增。简而言之,使用 ImmunoCult NK 细胞扩增包被材料(货号100-0714;STEMCELL Technologies)将未经组织培养处理的 12 孔板包被处理 2 小时。第 0 天,将重悬于 1 mL ImmunoCult NK 细胞扩增培养基(货号 100-0715;STEMCELL Technologies)中的 1 × 106 个 NK 细胞接种至包被处理的 12 孔板中。第 3 天,在每孔中加入 1 mL 新鲜的扩增培养基以培养细胞,第 7 天和第 10 天,将 NK 细胞重新接种到新鲜包被的孔板中,每孔 2 mL,密度为 0.2 × 106 个细胞/mL。NK 细胞共扩增 14 天。
人 NK 细胞的细胞因子过夜刺激
将人 NK 细胞与细胞因子混合液一起孵育,混合液包括 10 ng/mL 的 rhIL-12(货号 78027;STEMCELL Technologies)、20 ng/mL 的 rhIL-15(货号 78031;STEMCELL Technologies)和 100 ng/mL 的 rhIL-18(货号 592104;BioLegend,San Diego,CA)。这些均采用 ImmunoCult NK 细胞基础培养基制备,在 37 °C、5% CO2 条件下过夜孵育。
安捷伦 Seahorse XF 检测
将安捷伦 Seahorse XF HS PDL 细胞培养迷你板(安捷伦货号 103727-100)置于 37 °C、无 CO2 培养箱中过夜温育。对安捷伦 Seahorse XFp 探针板进行水化,并置于 37 °C、无 CO2 培养箱中过夜。在 XF 实验当天,使用 Agilent NovoCyte Advanteon 流式细胞仪测定NK 细胞总数和活细胞数量4。使用补充了 10 mmol/L 葡萄糖(安捷伦货号 103577-100)、2 mmol/L 谷氨酰胺(安捷伦货号 103579-100)和 1 mmol/L 丙酮酸钠(安捷伦货号 103578-100)的 XF RPMI pH 7.4 培养基(安捷伦货号 103576-100)制备安捷伦 Seahorse XF 检测液,并预热至 37 °C。将细胞悬液置于 15 mL 锥形管中并以 300 x g 离心 10 分钟。将细胞沉淀以 2.3 × 106 个活细胞/mL(第 0 天)或 1.2 × 106 个活细胞/mL(第 7–14 天)重悬于 XF 检测液中。使用 NovoCyte 流式细胞仪确认用于数据归一化的细胞浓度5。将 30 μL 的相应细胞悬液接种在预热的 XF HS PDL 迷你板(安捷伦货号 103727-100)中,每孔 7.0 × 104(第 0 天)或 3.5 × 104 个细胞(第 7–14 天)。接种后,将安捷伦 Seahorse HS 迷你板以 300 x g 离心 2 分钟。取下硅胶罩,并将孔体积调整至 200 µL(最终体积)。将微孔板置于无 CO2 培养箱中,在 37 °C 下孵育 45–60 分钟。采用 XF T 细胞代谢分析试剂盒(安捷伦货号 103771-100),并将来自安捷伦 Seahorse XF 细胞线粒体压力测试(安捷伦货号 103015-100)试剂的 FCCP 重悬于预热的 XF 检测液中。为了评估解偶联剂性能,将 BAM15 和 FCCP 以 30 µmol/L 重悬,并稀释至 25 µmol/L、20 µmol/L 和 15 µmol/L,以制备 10x 加药口原液。为了评估数天内的代谢情况,所有时间点使用的 BAM15 浓度均为 2.5 µmol/L(最终体积)6。
XF NK 细胞代谢分析检测的数据分析
安捷伦 Seahorse Analytics (SHA) 是一个用于分析 XF 结果文件的网页版软件平台。在 SHA 中,当使用 XF T 细胞代谢分析试剂盒试剂分析 NK 细胞时,XF NK 细胞代谢分析的专用分析视图可自动计算关键代谢参数(图 1)。
流式细胞分析
根据 BD Cytofix/Cytoperm Plus 方案(货号 555028;BD Biosciences,San Jose,CA),将人 NK 细胞与 BD Golgi Plug 试剂(含布雷菲德菌素 A)在 37 °C、5% CO2 条件下孵育 4 小时。使用 DPBS (Gibco,Billings,MT) 清洗 NK 细胞,使用 Zombie Aqua Fixable Viability 试剂盒(货号 423101;BioLegend,San Diego,CA)在室温下以 1/1000 的稀释比将死细胞染色 15 分钟,并在 FACS 缓冲液中清洗。加入 TrueStain FcX(货号 422302;BioLegend)封闭 Fc 受体,并置于冰上孵育 10 分钟。加入 anti-CD3、anti-CD56 和 anti-NKG2D 的混合液,以及相应的 FMOs,并在冰上孵育 20 分钟。使用 FACS 缓冲液清洗 3 次后,在冰上使用 BD Cytofix/Cytoperm 试剂固定细胞并进行破膜处理 20 分钟,然后使用 BD 破膜/清洗缓冲液清洗。在冰上使用抗人 IFN-γ 将固定和破膜的细胞染色 30 分钟,并使用 BD 破膜/清洗缓冲液清洗。利用 Agilent NovoCyte Advanteon 流式细胞仪和 Agilent NovoExpress 软件分析所有样品。所用抗体如下:抗人 CD3-FITC 抗体(货号 C344804;BioLegend)、抗人 CD56-PE (NCAM)(货号 318306;BioLegend)、CD314 (NKG2D) 单克隆抗体 PerCP-eFluor 710(货号 46-5878-42;Thermo Fisher Scientific,Carlsbad,CA)和抗人 IFN-γ 抗体(货号 502512;BioLegend)。
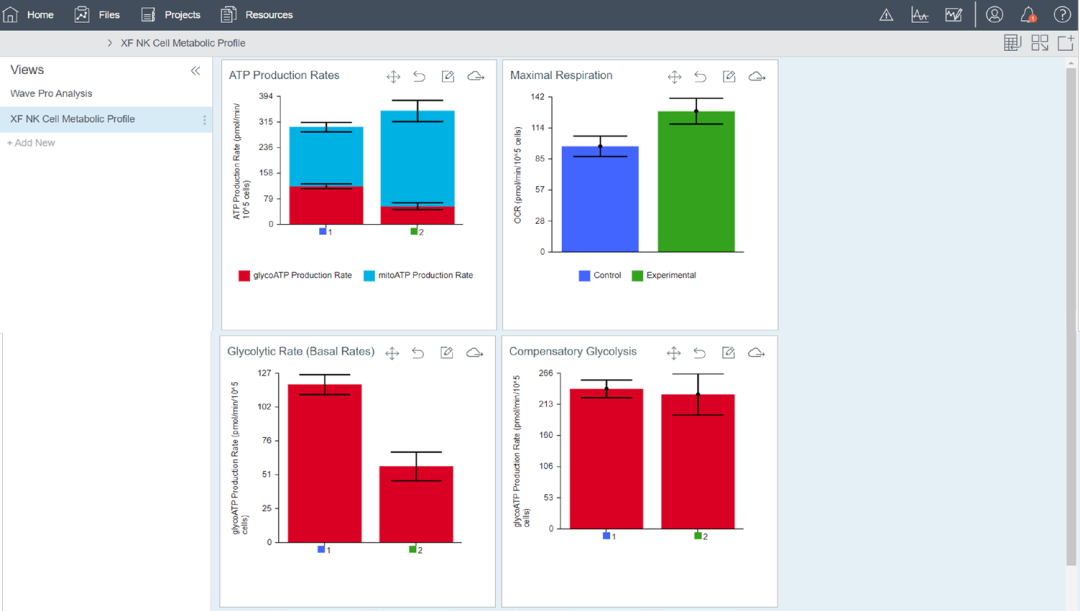
图 1. 用于 NK 细胞代谢分析检测的安捷伦 Seahorse Analytics 软件专用分析视图屏幕截图
结果与讨论
BAM15 解偶联剂在 NK 细胞中的活性评估
在评估活细胞的线粒体功能时,使用线粒体解偶联剂是一种常见的做法。使用含有解偶联剂 FCCP(羰基氰-对-(三氟甲氧基)苯腙)的安捷伦 Seahorse 细胞线粒体压力测试试剂盒,结合安捷伦 Seahorse XF 分析仪,可以提供有关免疫细胞能量代谢及其在指导细胞命运和功能中的作用的重要信息7,8。
在最近的研究中,我们证明了 FCCP 用于人或小鼠 T 细胞时无法提供最佳的线粒体解偶联性能,导致低估最大呼吸能力。
然而,含有解偶联剂 BAM15((2-氟/苯基){6-[(2-氟/苯基)氨基](1,2,5-恶二唑并[3,4-e]吡嗪-5-基)}胺)的 XF T 细胞代谢分析试剂盒可以更准确地测定最大呼吸。它为评估 T 细胞线粒体生物能量代谢能力提供了更强大的解决方案9。
因此,我们利用人和小鼠 NK 细胞评估了含有解偶联剂 BAM15 的 XF T 细胞代谢分析试剂盒试剂的性能,并将其与使用解偶联剂 FCCP(来自 Seahorse XF 细胞线粒体压力测试试剂盒)获得的代谢参数进行了比较。
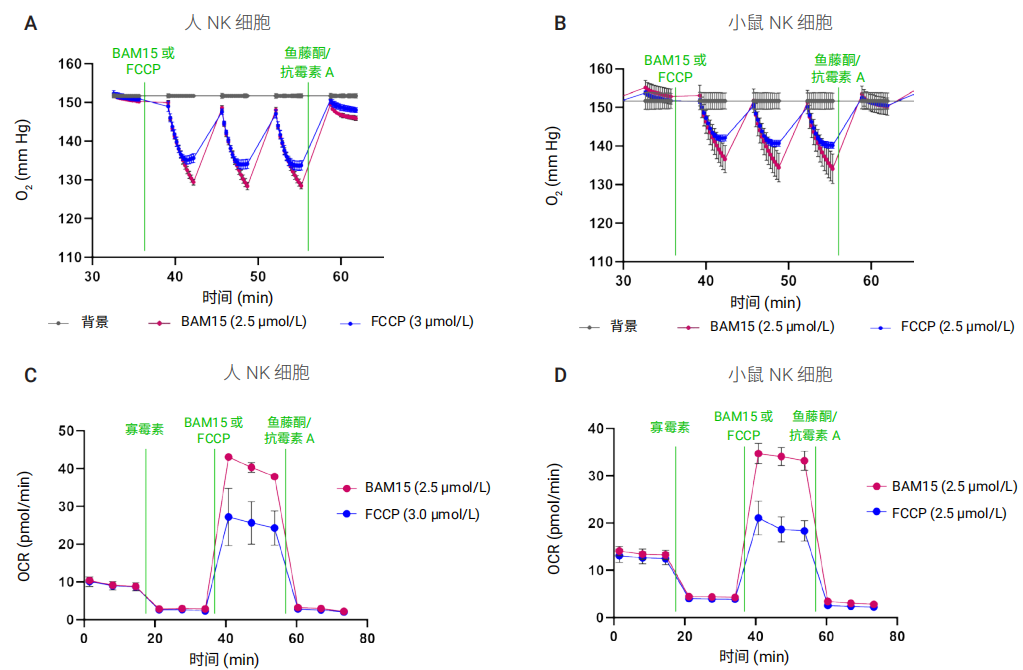
图 2. 比较使用解偶联剂 FCCP 和 BAM15 的耗氧量测量结果。添加最佳浓度的解偶联剂 FCCP(蓝色)和 BAM15(红色)后,来自 (A) PBMC 的人 NK 细胞和来自未免疫 C57BL/6 脾脏的 (B) 小鼠 NK 细胞中细胞外氧含量的变化。使用 T 细胞代谢分析试剂盒(红线)在线粒体压力测试(蓝线)或 NK 细胞代谢分析检测中从 (C) 人 NK 细胞和 (D) 小鼠 NK 细胞中获得的耗氧率 (OCR) 如图所示
如图 2A 到 B(蓝线)所示,使用 XF 细胞线粒体压力测试试剂盒检测人或小鼠 NK 细胞时,添加 FCCP 后,仪器测量的三分钟内细胞外 O2 浓度的变化速率并不是恒定或线性的。这导致了 OCR 计算的误差,并低估了最大呼吸(图 2C 和 D,蓝线)。而在使用解偶联剂 BAM15 替代 FCCP 后,添加 BAM15 后,仪器测量的三分钟内细胞外 O2 浓度稳定下降(图 2A 到 B,红线),实现了更准确的 OCR 和最大呼吸测定(图 2C 到 D,红线)。
为了证实与 FCCP 相比,BAM15 作为线粒体解偶联剂在 NK 细胞代谢分析中的性能更出色,我们使用来自至少三个不同供体或小鼠分离物的人和小鼠 NK 细胞进行了并行解偶联剂滴定。如图 3 所示,与在最佳 FCCP 浓度下获得的结果相比,以 BAM15 作为解偶联剂时获得的最大呼吸更高,标准偏差更低。滴定实验还表明,BAM15 的最佳浓度范围比 FCCP 的最佳浓度范围更宽。在所有条件下,对于同一供体使用 2.5 μmol/L BAM15 获得的最大 OCR 至少是在任何解偶联剂浓度下获得的最大 OCR 值的 90%,并且始终高于使用最佳 FCCP 浓度获得的值。这表明,当用于 NK 细胞时,BAM15 试剂解偶联性能优于 FCCP(图 4)。同时也表明使用 BAM15 时,极少需要针对每个样品优化解偶联剂浓度。
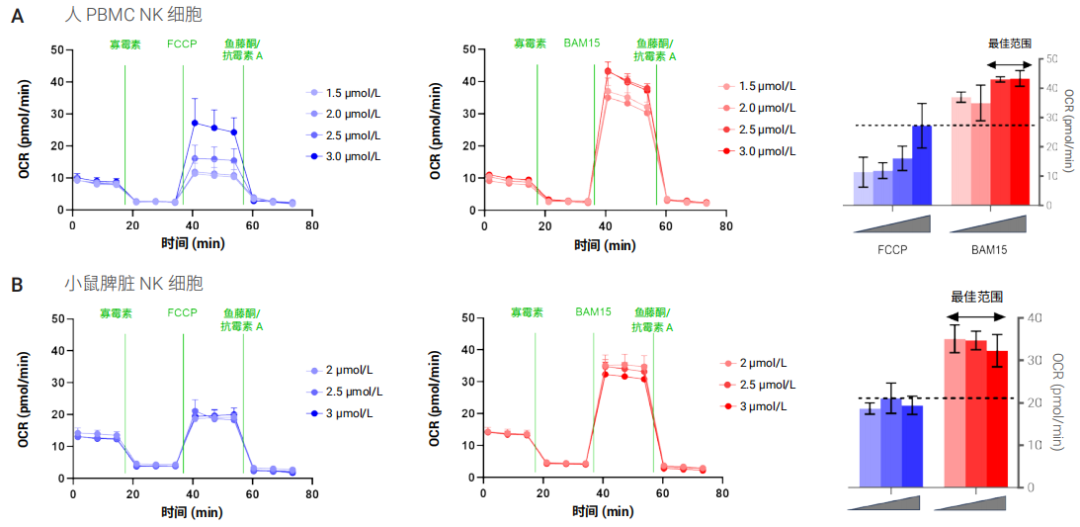
图 3. 比较 FCCP(蓝色)和 BAM15(红色)滴定实验中的最大 OCR 测量值。此图显示了使用 (A) 人 pbNK 细胞和 (B) 从未免疫 C57BL/6 小鼠脾脏中分离的 NK 细胞在不同解偶联剂浓度下获得的 OCR 动力学曲线和最大 OCR 的代表性数据。柱状图中的虚线表示在任意检测浓度下使用 FCCP 获得的最大 OCR
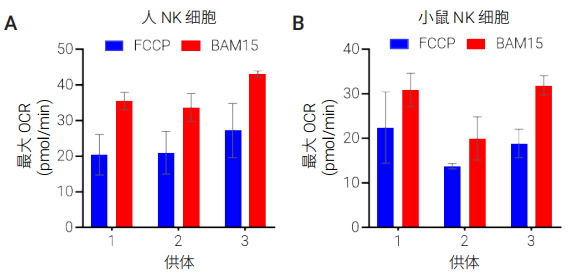
图 4. 比较在 (A) 人和 (B) 小鼠 NK 细胞中,添加最佳 FCCP 浓度时获得的最大 OCR 与添加 2.5 μmol/L BAM15 时获得的 OCR。数据表示三个重复孔的平均值 ±SD
利用 XF NK 细胞代谢分析检测方法监测 NK 细胞扩增过程中的代谢特征变化
糖酵解和 OXPHOS 在 NK 细胞功能的多个方面发挥着关键作用。两种代谢通路均不同程度地支持脱颗粒、Fas 配体 (FasL) 表达和干扰素-γ 生成,从而影响 NK 细胞的细胞毒性。例如,既往研究表明糖酵解对于 NK 细胞的细胞毒性、脱颗粒和 Fas 配体表达至关重要,而 OXPHOS 可能在支持 NK 细胞产生干扰素-γ 方面具有更主要的作用10。此外,糖酵解和 OXPHOS 可以相互影响,并协同支持 NK 细胞功能。例如,糖酵解可以为柠檬酸-苹果酸穿梭 (CMS) 提供中间体,CMS 将还原当量从细胞质转移到线粒体,为 OXPHOS 提供能量11。
由于两种生物能量代谢通路在支持 NK 细胞活化、增殖和效应功能方面具有重要作用,调节 NK 细胞代谢可能是改善基于 NK 细胞的癌症免疫疗法疗效的一种潜在策略。早期研究表明,如果要改善 NK 细胞过继转移方法治疗癌症的有效性,基础能量产生、糖酵解能力和总体代谢平衡是需要调节的关键参数10。监测细胞表型并将表型与功能和结果相关联的能力对于增强 NK 细胞适应性和抗肿瘤功能至关重要。
开发经优化的体外扩增方案需要能够提高细胞产量,同时支持 NK 细胞功能并避免耗竭状态。最近的研究表明,NK 细胞在扩增过程中进行了代谢重编程,从而形成了具有代谢灵活性的 NK 细胞,并能在营养匮乏的肿瘤微环境中持续杀伤肿瘤12。经过证明,扩增过程中存在不同的细胞因子组合会影响 NK 细胞代谢,同时改善 NK 细胞适应性。例如,使用 IL-2 或 IL-12/15 刺激人 NK 细胞可上调糖酵解和 OXPHOS 水平,而 OXPHOS 水平升高对于 NK 细胞效应细胞毒性和 IFN-γ 的产生至关重要10。经 IL-12/15/18 预激活的 NK 细胞也称为细胞因子诱导的记忆样 (CIML) NK 细胞,由于其在宿主体内更持久,被视为癌症免疫治疗的有力工具。它们的效应功能也得到增强,代谢也会转向糖酵解13。NK 细胞衍生的癌症治疗的另一种方法是使用来自患者的 PBMCs,通过 IL-2/15/18 刺激来大规模扩增自体 NK 细胞。这些细胞具有更长的寿命、对 K562 细胞的高细胞毒性和高水平的活化分子,如 CD16 和 NKG2D14。研究证明,单独暴露于 IL-15 可改善代谢适应性,包括 NK 细胞利用营养物质的能力。依次使用表达 IL-21 的饲养细胞和表达 IL-15 的饲养细胞,可提高功能性和体外肿瘤控制能力15。在体外扩增过程中,特定药物(例如,不同的细胞因子以及修饰或组合)可提供一种安全而有效的方法,用于建立对肿瘤微环境 (TME) 的不利代谢和免疫条件具有更强抵抗力的 NK 细胞群,例如细胞因子诱导的记忆样 NK 细胞,从而提高疗效16。然而,连续的体外刺激使 NK 细胞对细胞因子成瘾,当在无体内细胞因子支持的情况下输注这些细胞时,会导致其寿命缩短17。此外,细胞因子过度刺激 NK 细胞会损害NK 细胞功能,诱导代谢耗竭2。在扩增过程中监测 NK 细胞代谢情况对于优化 NK 细胞适应性至关重要,并可能有助于预测其功能命运。
XF NK 细胞代谢分析检测可全面表征 NK 细胞代谢特征,包括糖酵解和线粒体三磷酸腺苷 (ATP) 生成速率、代谢平衡、代偿糖酵解和最大线粒体生物能量代谢能力,所有参数均与 NK 细胞功能增强相关。
XF NK 细胞代谢分析检测使用之前经验证的用于 NK 细胞代谢分析的试剂和数据分析工具(安捷伦 Seahorse Analytics 软件),可快速提供 NK 细胞代谢表征的相关参数。
此处,使用 XF T 细胞代谢分析试剂盒(包括 NK 分析)监测 NK 细胞在扩增期间的代谢变化。在本研究中,使用市售扩增试剂盒 (STEMCELL Technologies) 扩增来自三个不同供体的人 pbNK 细胞。NK 细胞扩增使供体 1–3 的细胞数量分别增至原来的 10 倍、100 倍和 16 倍,在扩增 14 天内平均存活率为 70%–80%。在第 0、7、10 和 14 天收集扩增的 NK 细胞样品,并使用 XF NK 细胞代谢分析检测方法分析代谢特征。
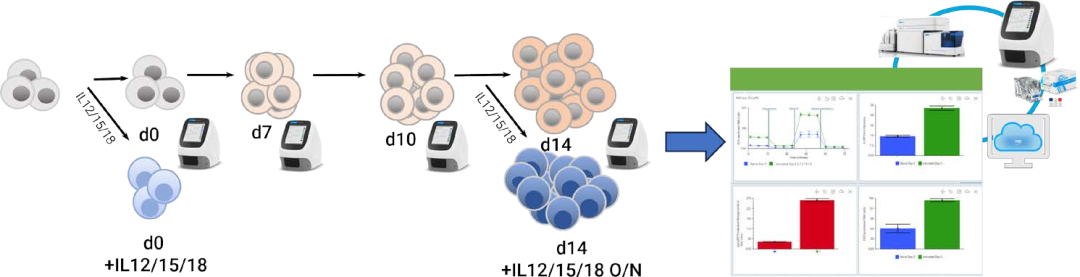
图 5.使用 ImmunoCut NK 扩增试剂盒 (STEMCELL Technologie),在扩增过程中评估人外周血 NK 细胞的实验设计示意图。在安捷伦 Seahorse XF HS Mini 分析仪上使用安捷伦 Seabgrse XET 细胞代谢分析试剂盒和 HSMini 细胞培养板进行 XFENK 代谢分析检测。使用安捷伦 Seahorse Anaics (SHA) 软件和 NK 细胞代谢分析的专用分析视图分析数据。在NK 细胞扩增前后以及使用 L12/15/18 刺激前后,分别使用 Agilent NovoCvte Advanteon 流式细胞仪对NK细胞开展流式分析
研究发现,NK 细胞的 ATP 生成速率随着扩增而提高,并伴随着代谢状态的变化。第 0 天,NK 细胞代谢需求较低(以 ATP 生成速率衡量),氧化代谢稳定,超过 80% 的 ATP 通过线粒体 OXPHOS 通路产生(图 6A)。在扩增过程中,两种生物能量代谢通路的活性均增加,但糖酵解活性增加更显著,占总 ATP 生成的 50%–70%(在第 0 天时为 10%-15%)(图 6B)。第 10 天,细胞达到最高基础代谢需求,线粒体 ATP 生成速率显著增加以满足这一需求。到第 14 天,基础代谢需求再次降低,30%–40% 的 ATP 生成来自糖酵解通路。在 NK 细胞扩增过程中糖酵解和 OXPHOS 活性均增加,但代谢特征从高度好氧(OXPHOS 依赖性)表型更多地转变为糖酵解表型。
第 14 天,用细胞因子 IL12/15/18 过夜刺激 NK 细胞导致所有供体糖酵解产生的基础 ATP 持续增加(图 6)。尽管不同供体之间的绝对代谢率不同,但细胞扩增过程中的代谢变化趋势一致。
NK 细胞代谢分析的另一个参数是代偿糖酵解,代表了当线粒体活性受到抑制时,细胞增加糖酵解 ATP 生成的能力。这一参数之前与 NK 细胞代谢潜力增加相关16。供体 1 和 3 的代偿糖酵解和最大呼吸均增加,但供体 2 未增加(图 6C 和 D)。
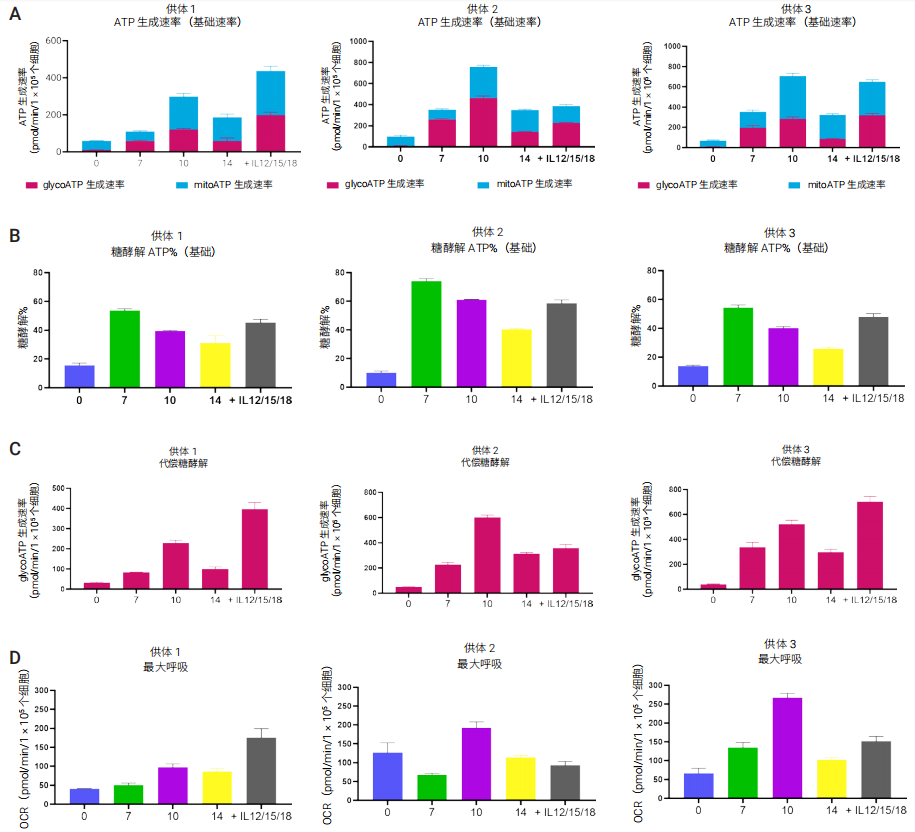
图 6. 来自三个不同供体的人 NK 细胞扩增和随后活化过程中的代谢测量。(A) ATP 生成速率,(B) 糖酵解 ATP%,(C) 代偿糖酵解,和 (D) 最大呼吸速率
NK 细胞对活化的反应程度因供体而异,这表明它们的成熟度和适应性存在差异。如果我们比较第 0 天(扩增前)和第 14 天(扩增后)的各个供体,会发现扩增细胞的代谢需求更高,糖酵解和线粒体 ATP 的速率都更高,如图 7 所示。
此外,对于扩增和未扩增的细胞群(第 0 天和第 14 天),使用 IL-12/15/18 过夜刺激均会导致糖酵解产生的 ATP 百分比增加。NK 细胞的总体代谢能力(以代偿糖酵解和最大呼吸衡量)也增加。然而,扩增的细胞达到了更高的代谢活性,具有更高的代偿糖酵解和最大呼吸。这说明这些细胞的代谢适应性增强,更有潜力满足额外的代谢需求。
由于 NKG2D-NKG2D 配体轴代表了人 NK 细胞介导的肿瘤细胞和病毒感染细胞识别的关键激活通路18,因此我们检测了激活受体在不同细胞群中的表达。在扩增的第 14 天,68% 的 NK 细胞表达 NKG2D 激活受体,而在第 0 天仅 4% 的细胞表达,表明细胞在扩增过程中被启动(图 8A)。
使用 IL12/15/18 进行短暂的 18 小时过夜刺激未改变该受体的表达水平。NK 细胞扩增前后均能够在 IL12/15/18 刺激下产生 INF-γ。在 IL12/15/18 处理的 NK 细胞中,表达 INF-γ 的细胞百分比增加,扩增前从 0.6% 增加至 67%,扩增后从 1.6% 增加至 94%(图 8B)。
综上所述,扩增后的细胞具有更高的基础代谢率,且更倾向于糖酵解表型。它们还具有更高的代谢能力,最大呼吸和代偿糖酵解均增加,这表明它们有更高的生物能量储备来满足压力条件下的能量需求。此外,这些扩增的 NK 细胞表现出更高的激活受体表达水平。当使用细胞因子过夜刺激时,扩增和未扩增的细胞的代谢率代偿糖酵解均进一步增加,同时产生更多的 INF-γ。所得结果与之前的研究一致,表明扩增方案可增强 NK 细胞代谢适应性、增加细胞毒性、上调激活受体水平,并增加葡萄糖和其他营养转运蛋白含量12–16。
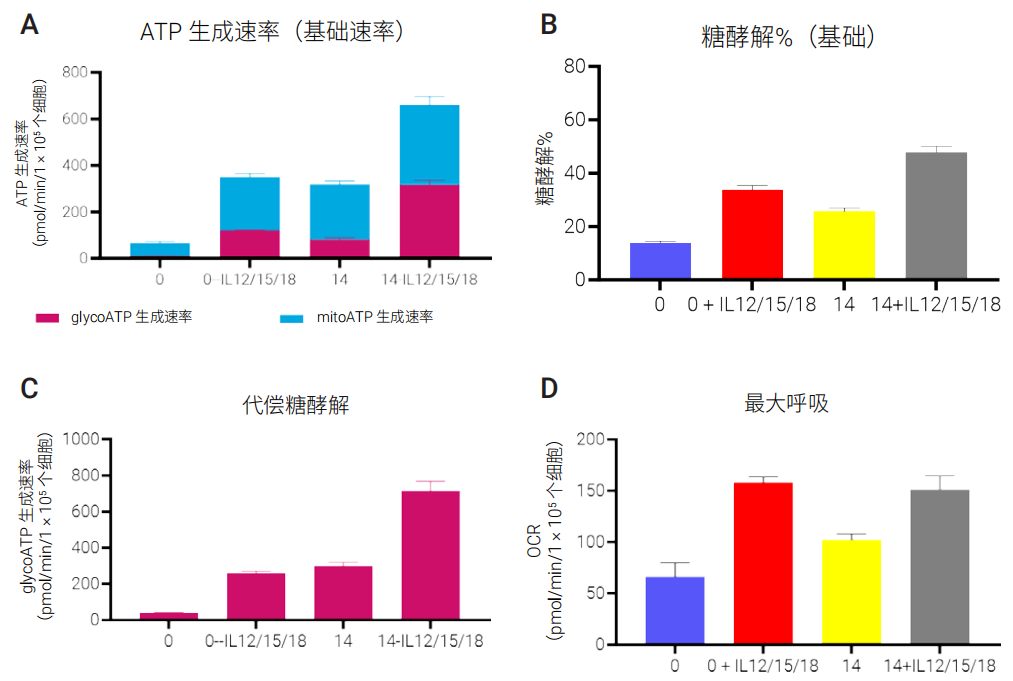
图 7. 供体 3 在 (A) 第 0 天、(B) IL12/15/18 激活后第 0 天、(C) 扩增第 14 天和 (D) IL12/15/18 激活后第 14 天的 NK 代谢特征
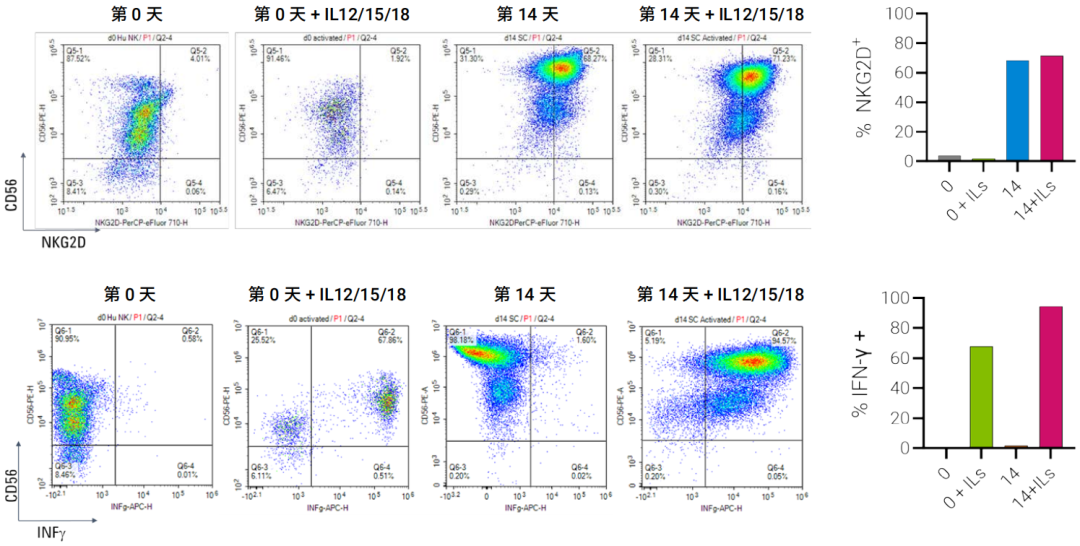
图 8. NK CD3-CD56+ 细胞在第 0 天、IL12/15/18 (ILs) 激活后第 0 天、扩增第 14 天和 IL12/15/18 激活后第 14 天的流式细胞分析。表达 NKG2D 和 INF γ 的细胞百分比
结论
安捷伦 Seahorse XF NK 细胞代谢分析检测可以全面表征 NK 细胞的代谢特征。该方法结合了安捷伦 Seahorse XF HS Mini 分析仪和 XF T 细胞代谢分析试剂盒,提供了优化的解偶联剂,能够可靠测量 NK 细胞的最大呼吸和备用呼吸能力百分比 (SRC),且仅需极少的解偶联剂浓度优化。此外,该检测方法使用户能够获得同一细胞样品的基础糖酵解和代偿糖酵解的定量信息。它还能够测量基础细胞生物能量代谢平衡和需求,无需进行多项检测即可全面表征 NK 细胞代谢适应性。
NK 细胞的代谢特征是提高 NK 细胞产品的持久性和抗肿瘤效力所需要分析的关键属性之一。更好地了解代谢和代谢底物的可用性如何影响 NK 细胞功能对于加速 NK 细胞相关疗法的设计和开发至关重要。XF NK 细胞代谢分析检测提供了多参数输出,提供来自同一样品的 NK 细胞代谢属性的详细数据。它可以作为常规检测方法纳入研究中,以优化 NK 细胞相关治疗产品的设计和生产。
更多详细内容
扫码下载该应用方案查看

参考文献
1.O'Brien, K.; Finlay, D.lmmunometabolism and natural killercells responses.Nature Reviews,2019, 19,282-290.
2.Laskowski, T.; Biederstadt, A.; Rezvani, K. Natural killer cellsin antitumor adoptive cell immunotherapy.Nat Rev Cancer,2022,22(10):557-575.
3.Gang, M; Marin, N.D.; Wong, P; Neal, C.C; Marsala, L.,Forster, M.; Schappe, T; Meng, W; Tran, J.; SchaettlerM.; et al. CAR-modified memory-like NK cells exhibitpotent responses to NK-resistant lymphomas.Blood2020,136(20),2308-2318.
4.Absolute Counting Using an Agilent NovoCyte FlowCytometer(使用 Agilent NovoCyte 流式细胞仪进行绝对计数),安捷伦科技有限公司,技术概述,出版号5994-6350EN,2023
5.Wang, G.; Pillai R.; Romero, N. Using the Agilent NovoCyteFlow Cytometer for lmmune Suspension Normalization inAgilent Seahorse XF Assays(在安捷伦 Seahorse XF 实验中使用 Agilent NovoCyte 流式细胞仪进行免疫细胞悬液归一化),安捷伦科技有限公司,应用简报,出版号 5994-6245EN,2023
6.Agilent Seahorse XFp T Cell Metabolic Profiling Kit(安捷伦 Seahorse xFpT细胞代谢分析试剂盒),安捷伦科技有限公司,用户指南,出版号 5994-3494EN修订版 B0,2022
7.Van der Windt, G.J.W; Everts, B; Chang, C.; Curtis, JD.; Freitas,T C; Amiel, E; Pearce, E.J.Pearce, E.L.Mitochondrial Respiratory Capacity is a Critical Regulatorof CD8+T Cell Memory Development.Immunity, 2012, 36,68-78.
8.Voss, K.; Hong, H.S; Bader, J.E.; Sugiura, A.; Lyssiotis, C. A.Rathmell, y.C.A guide to interrogating immunometabolism.Nature Reviews lmmunology,2021,21,637-652.
9.Walls J.; Romero,N.利用实时细胞能量代谢分析仪评估T细胞的生物能量代谢平衡和 SRC,安捷伦科技有限公司,应用简报,出版号 5994-4494ZHCN,2022
10. Wang,Z.; Guan, D.; et al. Glycolysis and OxidativePhosphorylation Play Critical Roles in Natural Killer CelReceptor-Mediated Natural Killer Cell Functions.Frontiersin lmmunology, 2020, 11, Article 202
11. Assmann, N.; o’Brien, K.L.; Donnelly, R. P; Dyck, L.;Zaiatz-Bittencourt, V; Loftus, R. M.; Heinrich, P; OefnerP J; Lynch, L; Gardiner, C.M.; et al. Srebp-controlledglucose metabolism is essential for NK cell functionaresponses.Nature lmmunology, 2017,18,1197-1206.
12. Poznanski, S.M; Singh, K. Ritchie, T. M.; Aguiar, J.A.,Fan, 1. Y; Portillo, A. L.; Rojas, E.A; Vahedi, F; El-SayesA.; Xing, S.; et al. Metabolic flexibility determines humanNK cell functional fate in the tumor microenvironmentCell Metabolism,2021,33, 1205-1220
13. Terrén, l; Orrantia, A; Mosteiro, A; Vitallé, y.;Zenarruzabeitia, 0; Borrego, F. Metabolic changes ofInterleukin-12/15/18-stimulated human NK cells.ScRep.2021,11(1)6472.
14. Liu, M., Meng, Y., et al. High-efficient generation of naturalkiller cells from peripheral blood with preferable cellvitality and enhanced cytotoxicity by combination of lL-2,lL-15 and lL-18.Biochemical and Biophysical ResearchCommunications, 2021, 534, 149-156
15. Zhang,C; Kadu, S; Xiao, Y;Johnson, O.; Kelly, A;O’Connor, R. S; Lai, M.; Kong, H; Srivatsa, S; Tai,V; et al. Sequential Exposure to lL21 and lL15 DuringHuman Natural Killer Cell Expansion Optimizes Yield andFunction.Cancerlmmunol Res., 2023, 11(11):1524-1537
16. Li, L., Mohanty, V, et al. Loss of metabolic fitness drivestumor resistance after CAR-NK cell therapy and can beovercome by cytokine engineering.Science Advances2023,9, eadd6997.
17. Miller, J.S. Therapeutic applications: natural killer cells inthe clinic.Hematology, 2013, 247-253.
18. Liu, H.; Wang, S; Xin, j.; Wang, j; Yao, C; Zhang, Z Roleof NKG2D and its ligands in cancer immunotherapy.AmJ Cancer Res., 2019,9(10):2064-2078.D01: 2156-6976/ajcr0099953